It has been a long time since my last visit to do some ringing in Laura’s garden, eight months in fact. Like Blakehill Farm, you need a wind free day, and they have been very few and far between. Laura collected the nets, poles and other equipment on Monday to give her the opportunity to practice putting up the nets, furling them and securing them so that they don’t come undone overnight, as I am putting her forward for her C-permit this Spring. She, no doubt helped by Adam and Daniel, did an excellent job, with the added bonus that I could have a lie in and didn’t have to do any of the setting up.
Unfortunately, the flocks of Goldfinch, sometimes 50 strong, must have felt that something wasn’t quite right and they didn’t flock at all. The usual Blue and Great Tits arrived, including this rather sooty / dusty looking individual:
Blue Tit, Cyanistes caeruleus
We did eventually catch a few Goldfinch, but the bird of the day was a female Great Spotted Woodpecker. It can be difficult to age them at this time of year, as they moult in strange ways. However, this one’s black plumage was just that, all black and glossy. Definitely a full adult: I meant to take a photo but it was released before I blurted out that I wanted to do so. Looking at my stock photos made me realise that I only have one of an adult female, must make sure I get some more.
The catch for the session was: Great Spotted Woodpecker 1; Blue Tit 15(2); Great Tit 4(1); Long-tailed Tit 2; Goldfinch 3. Totals: 25 birds ringed from 5 species and 3 birds retrapped from 2 species, making 28 birds processed from 5 species.
Not our greatest haul but one of the benefits of garden ringing is that it is somewhat more comfortable and relaxing, with coffee and biscuits on tap. Besides, the way the weather is shaping up we will be lucky to get more than four sessions in this month. Yesterday’s planned session had to be abandoned because, although the forecast morning rain did not arrive until the afternoon, the light winds turned out to be strong winds gusting up to 35mph, far too windy for setting nets and, coming from the East, bitterly cold. Grateful for small mercies! Fingers crossed for Saturday.
We were a little unlucky: whilst opening the nets a Goldcrest put in an appearance but decided not to hang around to get caught and Mr and Mrs Blackbird were happily hunting on the ground but just kept flying under the nets, until we lowered them almost to the ground, whereupon the pushed off. Oh well!
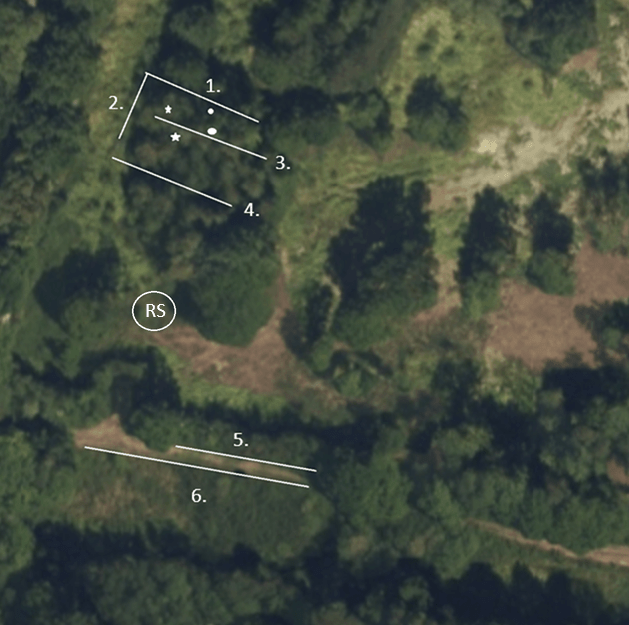
Our first opportunity to get out for ten whole days! A big relief. I got out in a light shower yesterday to top up the feeders for today’s session at Somerford Common. I was joined for the morning by David, Laura, Adam, Pete and Claire. We arrived on site at 7:30, having had to defrost our vehicles and to a temperature of -1oC. Thankfully with a clear blue sky, and very little cloud anywhere, it soon warmed up and we set the usual nets:
Once upon a time, not that long ago, rides 5 and 6 were key to a good catch. It was where we would catch Redwing, Goldcrest and Long-tailed Tit. All three of these have massively reduced in our catch at this site. Today those five nets produced a solitary retrapped Blackbird! There is no obvious reason why that should be the case but they have been very poor all winter.
Continuing the good start to the year, our first round produced three new and one retrap Marsh Tit. We caught another two retraps later in the session. In the first six weeks of this year, and in just six sessions, we have ringed seven and retrapped 17 Marsh Tits: looking good for another decent year for the species in the Braydon Forest.
We caught and ringed another two Chaffinch this session, but also had to let another three go: one had a mite infection, the other two had Fringilla papillomavirus.
The list for the session was: Nuthatch 1(3); Blue Tit 13(17); Great Tit 3(8); Coal Tit (7); Marsh Tit 3(3); Long-tailed Tit 1; Robin 1; Blackbird (1); Chaffinch 2. Totals: 24 birds ringed from 7 species and 39 birds retrapped from 6 species, making 63 birds processed from 9 species.
At 11:30, with the catch having died away, the breeze got up and we decided to close up and take down. Whilst doing so, Claire heard some bird calls she hadn’t heard before. When she described seeing a small group of five birds: one red and four greenish-yellow Pete played her the call of Crossbill, which she recognised straight away. They didn’t hang around for long but great to know they are about. I feel a session on the western side of Somerford Common coming soon: as that is where the main conifer blocks are. I will have to find a suitable pond! They are notoriously difficult to catch, as they stay in the treetops unless c0ming down to drink!
Anyway, we had a few more birds to process before packing away but with many hands making light work, it didn’t take long and we were away from site by 12:30. Well I would have been if Claire and I hadn’t gone Crossbill hunting!
We are going to try a few different net positions next time, alongside the treeline that lines the road down to our parking area. Forestry England have cleared the line within the paddock and we often see good numbers of birds, not Blue Tits thankfully, moving along that stretch.
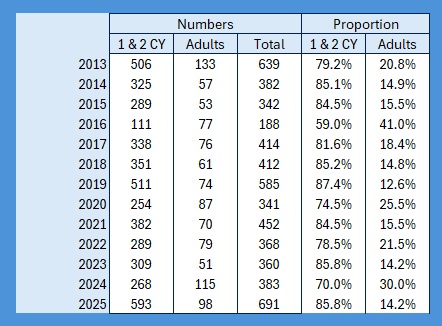
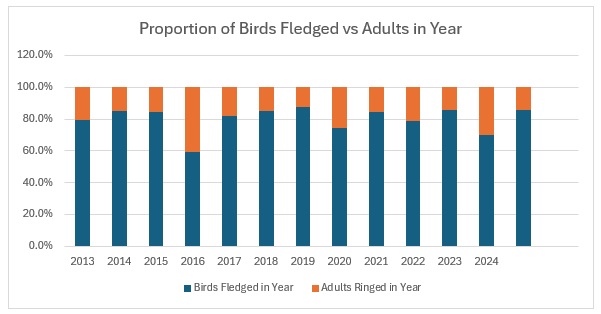
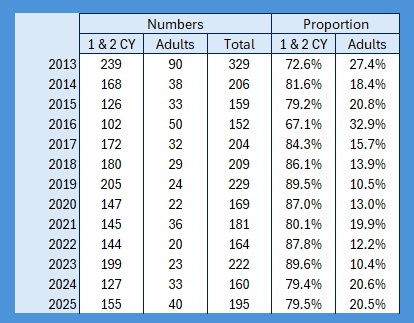
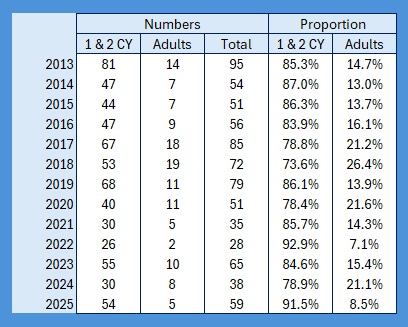
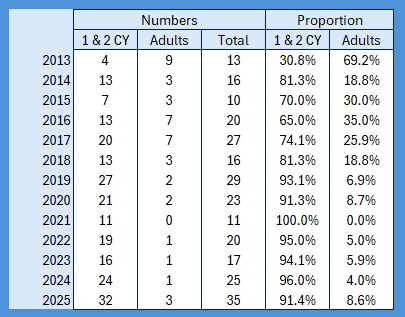
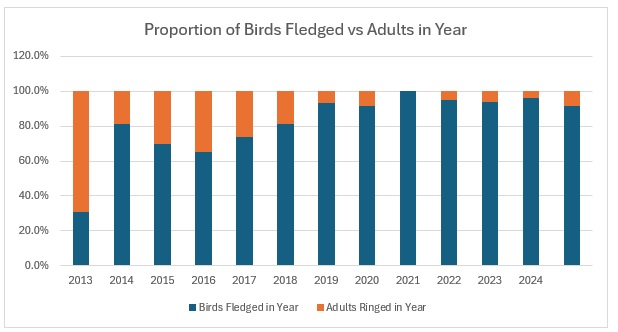
It’s raining, it’s pouring, no ringing, it’s boring! So I thought, having looked at the population levels of the Titmice being ringed in the Braydon Forest, whether these changes were reflected in the split between young and adults. To do this I counted all of the birds ringed with a BTO code of 3 and 3J for the year, and then I counted all of those ringed with a BTO code of 5 in the next year. For adults I counted all in the year with a BTO code of 2, 4 or 6. Not 100% accurate as a code 2 in a year means nobody knows when it fledged. This should only be an issue for species like Nuthatch, House Sparrow and Long-tailed Tit where both adults and juveniles moult into full adult plumage in the autumn. Everything code 2 becomes code 4 on the 1st January of the following year. , code 4 also applies to all birds, except codes 3 and 3J, once the breeding season is over and the post-breeding moult is under way. Code 6 means that they are definitely fully adult but, again, that reverts to code 4 after the breeding season. I have included figures for 2025, which is a little unfair as we will almost certainly catch more of each age group between now and June / July when the 2026 youngsters will fledge, so take those with a pinch of salt.
Blue Tit, Cyanistes caeruleus:
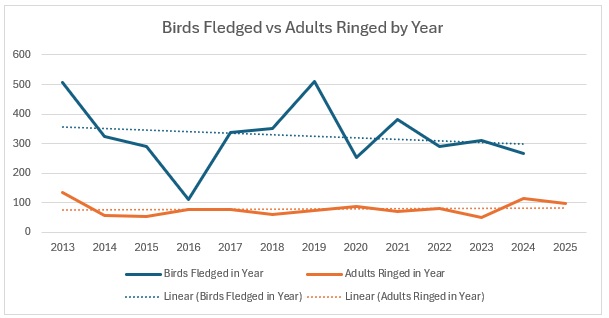
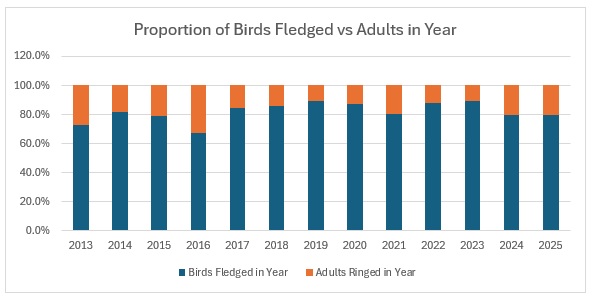
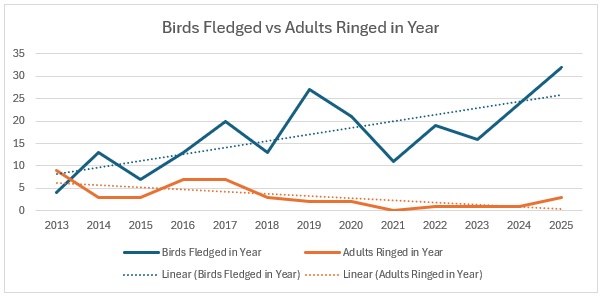
As the largest cohort of birds in the Forest, I started with Blue Tits. My previous post on their population in the Braydon Forest showed that it is relatively stable, but with a slow decline:
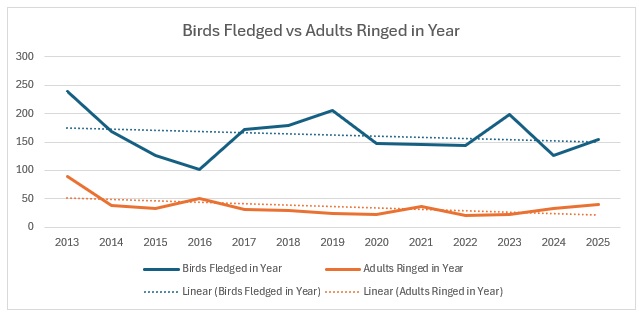
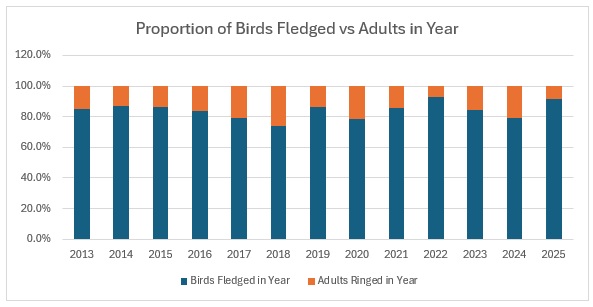
This shows clearly that 2016 was the worst breeding season for this species in the 13 years covered by the project. That shows in the stable trend line for adults but the decline in juvenile birds. When we look at the catch with juveniles vs adults as a proportion of the totals ringed we get this:
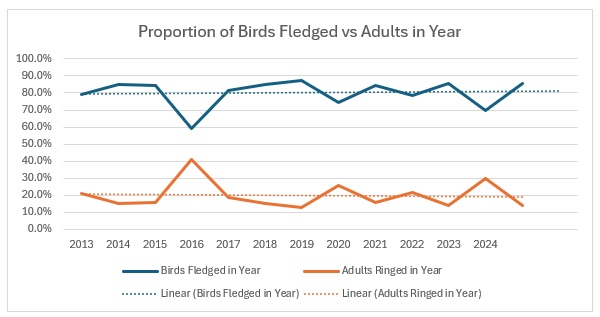
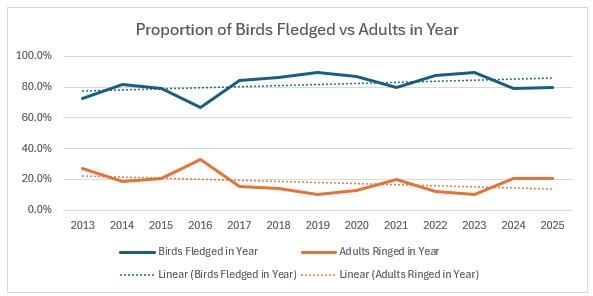
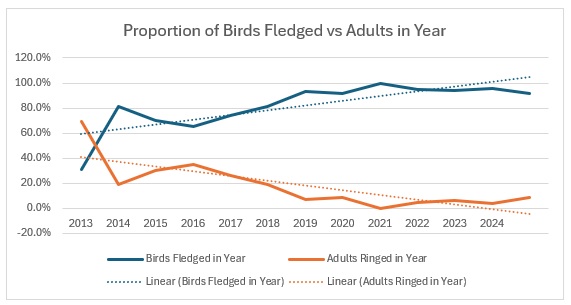
As you can see, across the entirety of the period, the trend in the catch is much more equivalent for both age groups than the numbers would indicate. The bar graph shows exactly the relationship between adults and juveniles:
The average split over the year is 80:20 and relatively stable, excepting 2016.
Great Tit, Parus major:
The previous post on the status of Great Tits in the Braydon Forest showed another species with something of a decline:
As you can see, both adult and juvenile numbers have declined in harmony with each other. When we look at the proportional analysis:
So, juveniles are increasing with respect to the number of adults being ringed. So, is the overall decline to do with a reduction in the number of adults surviving year on year?
As with the Blue Tit, the Great Tit clearly had a bad year in 2016, as the adults made up a higher than usual proportion of the total. Their average split though is 82:18, juvenile to adult.
Coal Tit, Periparus ater:
This species showed the biggest fall in [population over the period:
Looking at it in pure numbers, it does look as the key issue is adult survival. However, when you graph it up it is the juvenile numbers that show the bigger trend decline:
Proportionately, though:
Yet again, there is a slight increase in the proportion of juveniles to adults ringed across the period.
Again, as you can see, the proportions are largely juvenile birds. This time the ratio across the years is 84:16. Interesting to note that they were not affected in the same way as Blue and Great Tits in 2016, but did have a dip in the proportion of juveniles in 2018.
Marsh Tit, Poecile palustris:
Now to my key project species, to which I have already dedicated a lot of time, analysis and writing:
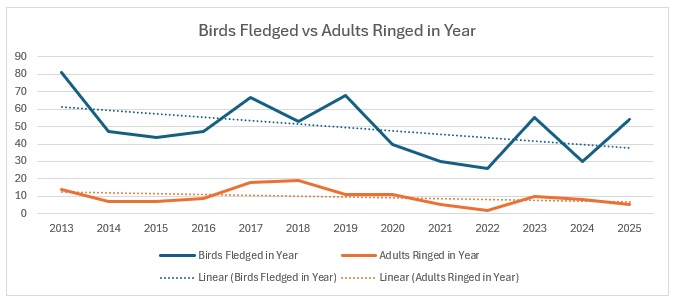
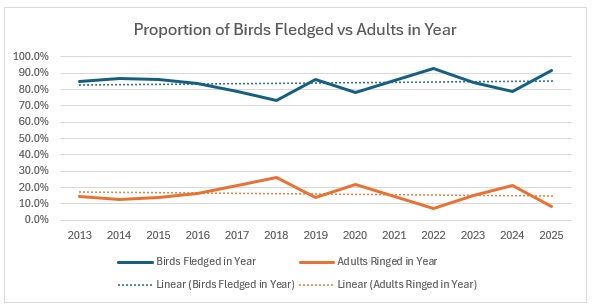
This is a real astonishingly different situation. Obviously, with a much smaller population, being a red-listed species, any changes are going to be amplified, so that needs to be borne in mind. However:
It is pretty clear what is happening here: juvenile survival is surprisingly strong, having started from a very low base, and adult survival is very much lower. Proportionately this is how it looks:
Obviously the concern is that, if we continue to have so few adults being caught, how long can the number of juveniles continue to increase?
I have absolutely no idea why the situation with Marsh Tits started with such a huge disparity between adults and juveniles in 2013. I hate to say it but perhaps my ageing capabilities for this species weren’t up to scratch in 2013, my first full year of solo ringing. (Then I purchased Jenni & Winkler: Moult and Ageing of European Passerines, the absolute bible of ageing, and have never looked back. First as a Kindle download and then the real thing when the second edition came out in 2020. Fabulous book.) Including that gives a proportion of 82:18. Excluding that year changes the proportion to 84:16. Obviously the unusual year was 2021: no adults ringed at all. I have never seen that in any of our regularly caught resident birds.
It is not that I am obsessed by titmice but they provide lots of data for looking at the health of our woodlands in supporting various bird populations.
With the rest of this week, and the start of next week, forecast to be wet and miserable it was lucky that today was forecast to be fine, after heavy rain all day yesterday. Sure enough, at 7:15 when I went to put the nets and things into the car, it was very wet on the ground but just a bit damp in the atmosphere but, by the time I got to site, the mist lifted and the humidity decreased to tolerable levels.
I was joined by Miranda and Ellie for the morning. We met up at 7:45 and set up the following nets:
Regular readers will notice that we did not set the nets along the track to the east of the ringing station. It is muddy at the best of times but, after the recent rains, it was an absolute quagmire and I felt we would be losing wellies every other step!
I topped up the feeders before we set the nets. I had planned to do it yesterday but the weather was so dreadful I decided against getting soaked. The first birds arrived at 9:00. As has become usual, the catch was Blue Tit heavy.
We didn’t have a stunner, like Monday’s Sparrowhawk, to relieve the constant pecking. A lovely male Chaffinch, a group of four Long-tailed Tits (one male and three females) and three retrapped Marsh Tits. It was a less pressured session than recent ones but we ended up with a reasonably decent haul: Blue Tit 24(11); Great Tit 2(1); Coal Tit 3(3); Marsh Tit (3); Long-tailed Tit 4; Robin 1; Chaffinch 1. Totals: 35 birds ringed from 6 species and 18 birds retrapped from 4 species, making 53 birds processed from 7 species.
The wind began to get up at 11:00, so we did a last round at 11:15, and closed the nets as we emptied them. We processed the remaining birds and started taking down and packing away. We left site at about 12:15.
I cancelled Saturday’s session in Ravensroost due to a weather forecast that said wet and windy all morning. Waking at 6:15 to find it was raining, I thought “back to bed”. When I woke again at 8:00 it was dry, bright and calm: that is, the weather was, I in a foul mood. Still, I had agreed to run a session for the Chippenham Men’s Mental Health Group on Monday and Ravensroost is the best place to carry that out, so we rescheduled for today.
I was joined for the session by Laura, David and Ellie. We arranged to meet at 7:45, as the weather was still forecasting rain until 7:00, having thrown it down pretty much all day Sunday. I set my alarm for 7:00. At 6:10 my good lady wife woke me up asking if I had cancelled the session. According to her clock it was 7:20. Cue panic from me! I dashed into the kitchen to find the clock showing 6:10! Something untoward had happened to her clock! I dozed for the next half-hour but no more sleep.
Fortunately, when I left the house at 7:30, although there was still a lot of damp in the air, it wasn’t raining. Of course, things could not run totally smoothly: I arrived at the site 10 minutes early, but spent the next 15 minutes trying to get the padlock open. Unlike every other Wiltshire Wildlife Trust site, somebody replaced the standard padlock, all opened by a single key, with a combination padlock. It has been a pain since it was installed: frequently seizing up. In the summer it was sap or syrup from some bushes and fruits, this winter goodness knows. Eventually some liberal spraying of WD40 and some brute force with a pair of pliers did the trick! I understand the Trust are going to replace it with a standard Padlock: the sooner the better!
Anyway, I had filled the feeders on Friday, in anticipation of Saturday’s session. They were empty on arrival so we refilled them. It did, of course, indicate that we would be enjoying the attention of Blue and Great Tits. As you can probably guess from the title. we were not “disappointed”! However, the first bird out of the nets was a titmouse: a new Marsh Tit, our fourth already this year.
We set our usual nets and waited for the deluge: we didn’t have to wait long. It was a very busy morning, so I was pleased that there were the four of us to manage the load.
We had some very decent birds in the catch, the best of the lot was this beauty:
Juvenile male Sparrowhawk, Accipiter nisus, thanks to Laura for the photo and for blurring me out (I don’t like having my face in photos)
The Chippenham group arrived at 10:45, just in time to see the Sparrowhawk processed. Because none of my team, apart from me, had ever ringed a Sparrowhawk, I got them to draw lots for it: Laura won and had the wonderful experience of being spiked by those fabulous talons! They are their killing tool and require careful handling. We sexed it based on the biometric data: females are considerably larger than males. So much so that the females take a larger ring size, so it is important to get the sexing correct!
As well as this, we ringed our first two Dunnock of the year, and retrapped our third.
The Chippenham crew were good company, and very interested in the details of the ringing scheme. We managed to show them how we catch and extract a variety of birds, together with every bit of the process. As we had a large number of birds to process, after I thought they had seen all of the potential variety, the group leaders and I suggested they head off for a walk, so they didn’t get bored. We then cracked on with processing the last forty birds. Fortunately, David’s dad, Trevor, had turned up to help us pack up and take David home afterwards. He very kindly took on a lot of the scribing: doing B-rings and retraps, whilst I scribed A-rings and processed a chunk of the remaining.
The list for the day was: Sparrowhawk 1; Nuthatch 1; Blue Tit 45(14); Great Tit 8(13); Coal Tit 1; Marsh Tit 1(2); Long-tailed Tit 1(1); Wren 1; Dunnock 2(1); Robin 3. Totals: 64 birds ringed from 10 species and 31 birds retrapped from 5 species, making 95 birds processed from 10 species.
We shut the nets whilst doing the last round, so we didn’t catch any more birds. By the time we had processed that lot it was gone midday. However, we split into two teams and got the nets down quickly. We were off-site by 13:00 after a busy Blue Tit heavy session.
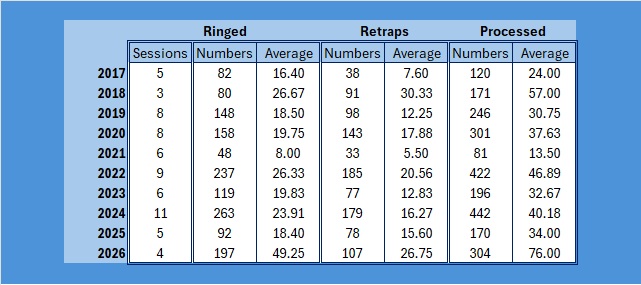
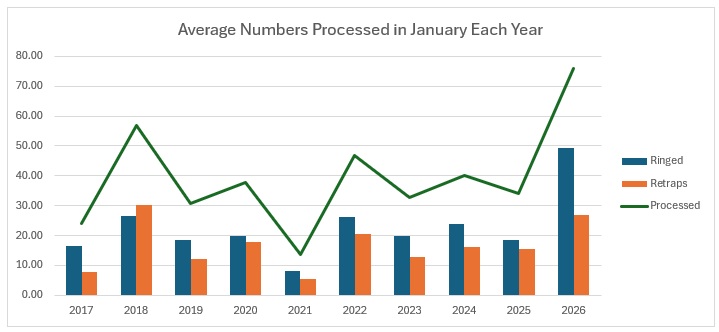
A quite remarkable month given how awful the weather has been. To put it into perspective, my team only managed four sessions, but we processed 304 birds from 16 species. 50% Blue Tit but good practice for the team!
This is the catch:
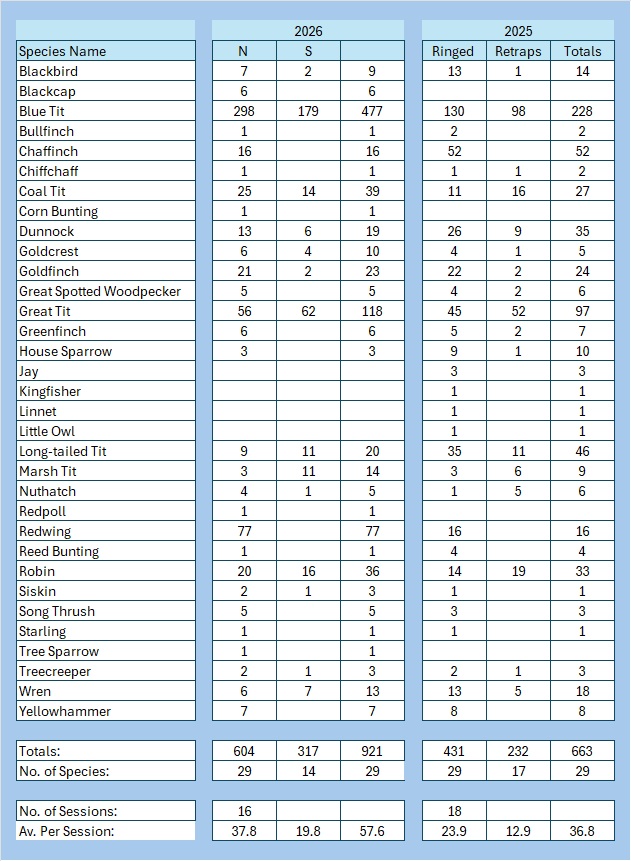
So the Group as a whole only managed 16 full sessions and we processed 921 birds from 29 species, compared to 663 from 29 last January. Ironically, we processed four species that we didn’t last year and last year we processed four species that we didn’t catch this January. We had a bigger catch of 1,241 from 33 species in 2024, but that came from 34 sessions.
Added to this January’s list were Blackcap, Corn Bunting, Redpoll and Tree Sparrow. Missing from this January’s list were Jay, Kingfisher, Linnet and Little Owl. Of course, it would have been lovely to have had those on our list this year as well.
On the whole, the key difference is the astonishing number of Blue Tits ringed: over 160 more than last year. It is far and away our largest catch of them by session than any other January. 80% of those ringed were second calendar year birds, the average is 73%. They have clearly had a good breeding season with, so far, a good survival rate over the winter to date. Other species showing improvement in numbers over last year were Coal Tit (39 vs 27) and Redwing (77 vs 16). However, there were significant declines in the catch of more species: Chaffinch (52 vs 16), Dunnock (35 vs 19 caught but, significantly, 26 ringed vs 13), Long-tailed Tit (both had 11 retrapped but ringed was 35 vs 9).
Let’s hope for an improvement in the weather in February
January has been a very difficult month. I had planned for ten sessions this month. Thanks to the weather and the forecast for this coming Saturday, it looks as if we will only have managed four of those sessions! I will return to this at the end of the session report.
I was joined for the session at 7:30 by Laura and Pete and we set the nets down the central glade. However, with Pete needing to leave for work at 10:30, and there only being the three of us, I decided to only set the nets from the bottom of the hill onwards, so we had 3 x 16m and 5 x 18m set up and open by 8:15.
Of course, every session this month has delivered firsts for the year. Today’s were: Redpoll, Blackbird and Song Thrush. This was the first bird out of the nets this morning:
2nd Year Song Thrush, Turdus philomelos
It flew into the net before we had a chance to open it. Fortunately, it just sat in the pocket of the top shelf and didn’t wrap itself in lots of net. In fact, I was just able to reach in and lift it out. That Song Thrush is the first that my team have captured and ringed since the 20th August 2025! The Redpoll was the first we have caught since the 20th November 2025 but we caught our previous Blackbird on the 31st December 2025.
The weather was dry with no wind, but that meant that we spent the entire morning shrouded in mist. The sun threatened to break through for about half-an-hour at 10:00, but then disappeared That also has an impact on bird movements and on the nets. They become beaded with moisture, making them more visible, and incredibly wet by the end of the session. My nets from this morning are currently hanging in front of the radiator in the bathroom in the hope they will dry out before I need them again.
The list from this morning was: Blue Tit 14(14); Great Tit (3); Coal Tit (3); Long-tailed Tit (1); Wren (1); Robin 2; Redwing 2; Song Thrush 1; Blackbird 1; Goldcrest (1); Chaffinch 3; Redpoll 1. Totals: 24 birds ringed from 7 species and 23 birds retrapped from 6 species, making 47 species processed from 12 species.
It never warmed up at all this morning and, after Pete left at 10:30, Laura and I agreed to pack up at 11:30. We did our last round then and closed up the nets. There were only a couple of birds, so we left them in the bags whilst we took the nets down: it only took us 15 minutes to get them down, and then we processed the remaining birds. The benefit of that was one less trudge up that hill! It really is the most exercise we get at any of our sites. I need a stick to help propel me up it these days! We had it all packed away and were off site by midday!
To return to the theme that I opened with, it looks as if Saturday is going to be horribly wet, so I think that four sessions is all we are going to get this month. Quite remarkably we have processed 304 birds from 16 species, all in local woodlands of the Braydon Forest – mainly we were restricted to woodlands because our other sites were too exposed to the weather.
As you can see, it is not our largest catch in January, but it is our highest average number processed, our highest ever average ringed but not our highest ever retrap average. All I hope is that February is a lot better weatherwise.
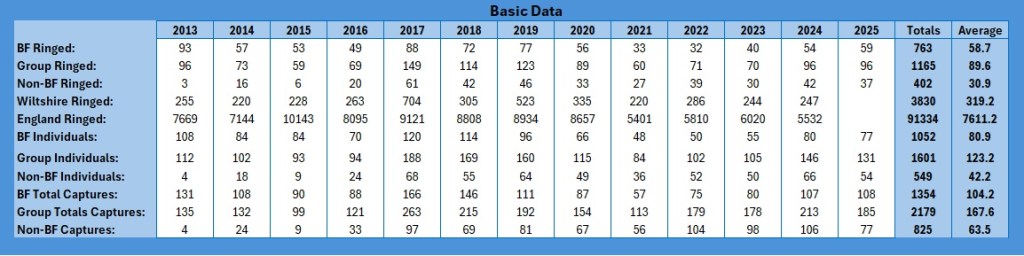
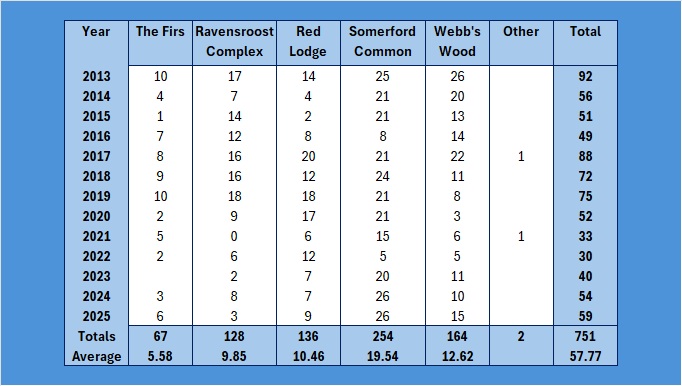
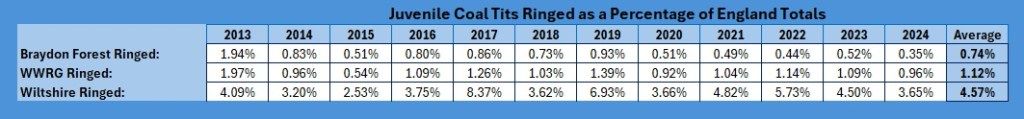
So, a final analysis in my four part Paridae examination within the Group, the subset of the Braydon Forest, and how they compare with the numbers for Wiltshire and England as a whole. This time it is our lovely Coal Tits, Periparus ater.
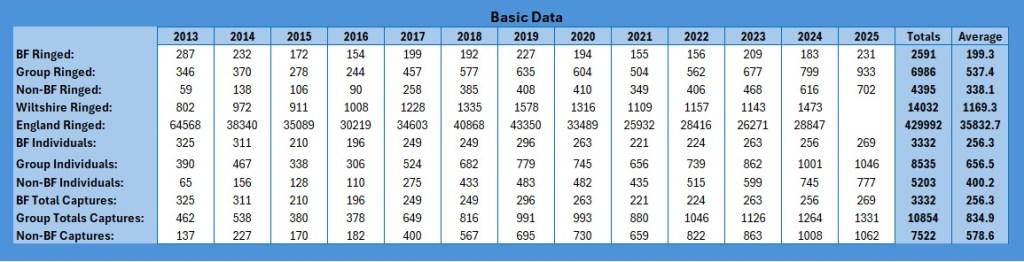
So, to start with, these are the basic figures:
As with the other data sets, I have restricted the graphs to 2024 because the data for 2025 has not been released yet. So how do these basic figures look graphically?
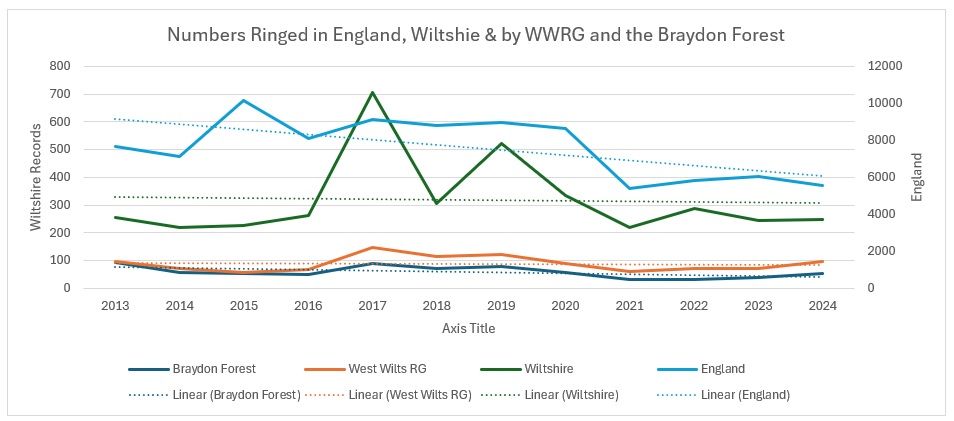
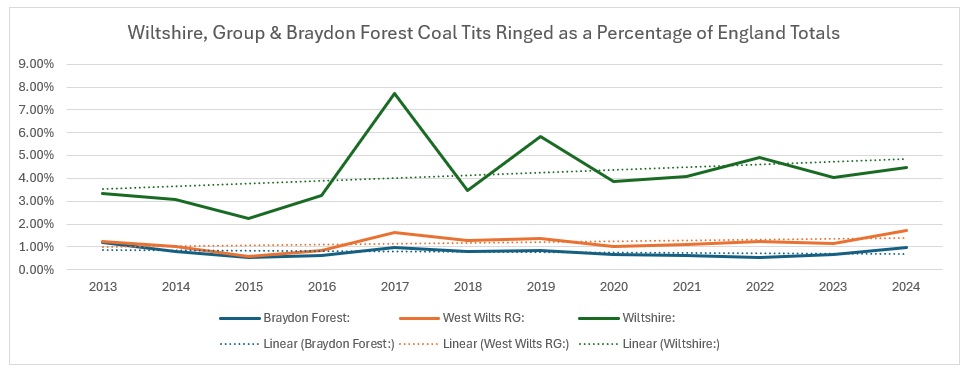
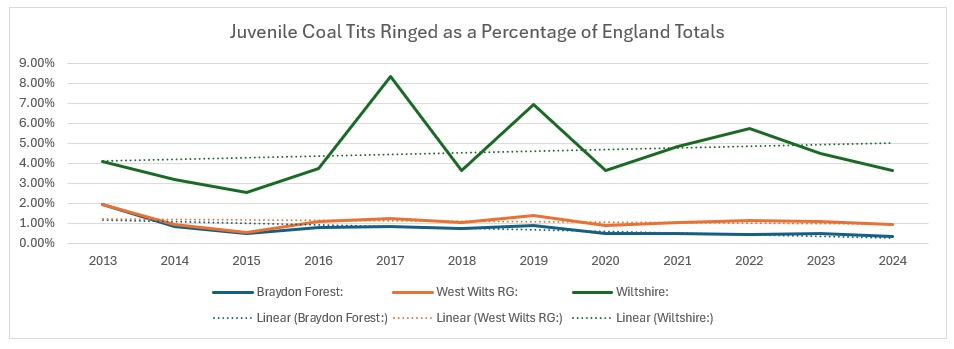
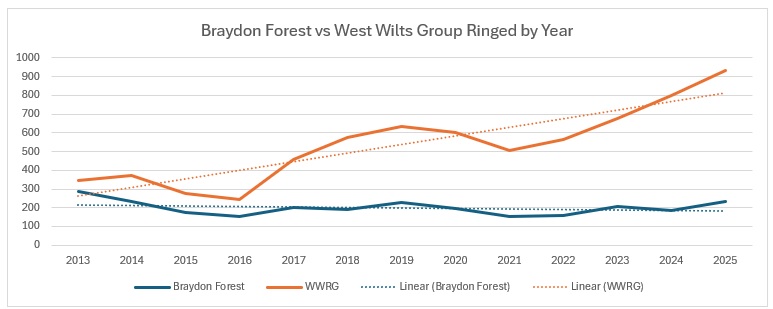
As you can see from the trendlines, the decrease across England is not as pronounced as it is across Wiltshire and within our group areas. As a proportion of the England catch it looks like this:
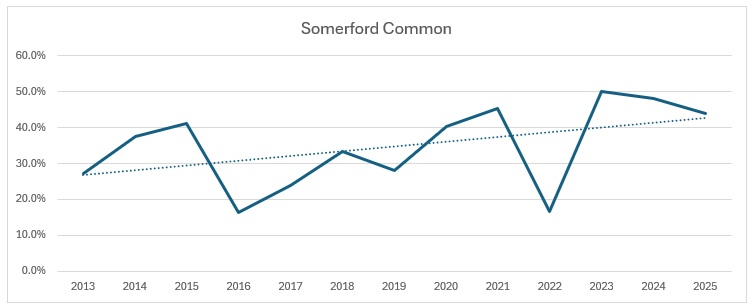
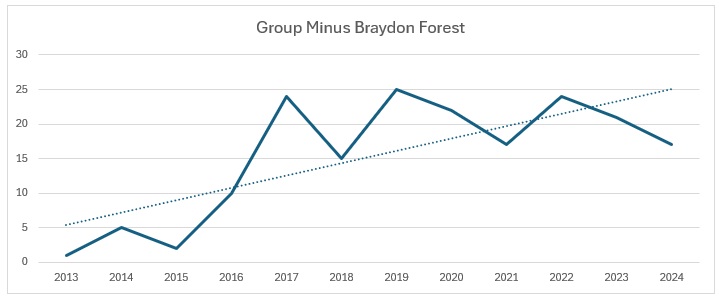
It is pretty clear that Wiltshire as a whole is bucking the national trend, whereas our group results show a slight increase, whilst the Braydon Forest is showing a slight decline. I am not sure why: the main site for them in the Forest is the Somerford Common complex and it is the one area that has undergone the least change over the 12 years of the study. Unlike the other four species, I decided to have a look how they split across the Braydon Forest:
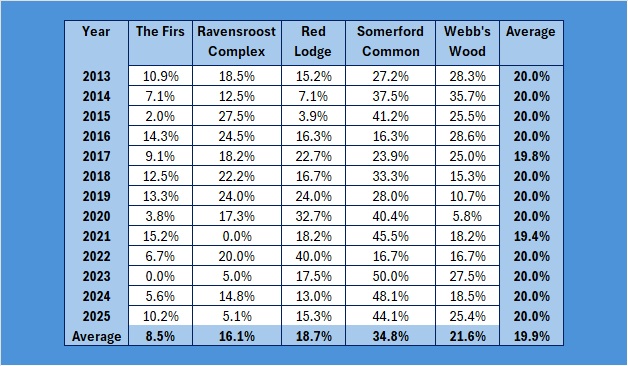
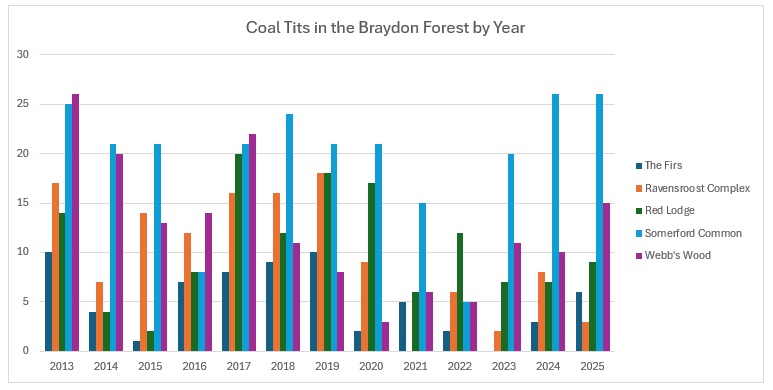
Other is my back garden! As you can see, Somerford Common has the highest number caught over the years. Proportionately, it looks like this:
If you graph that up, it looks like this:
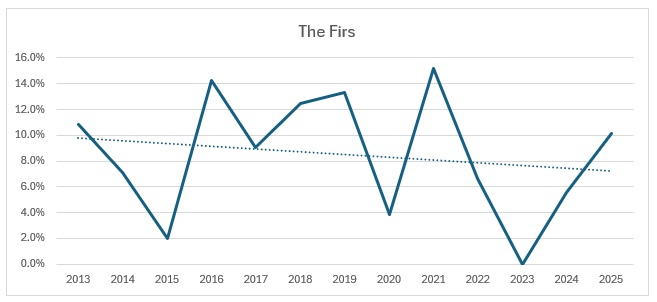
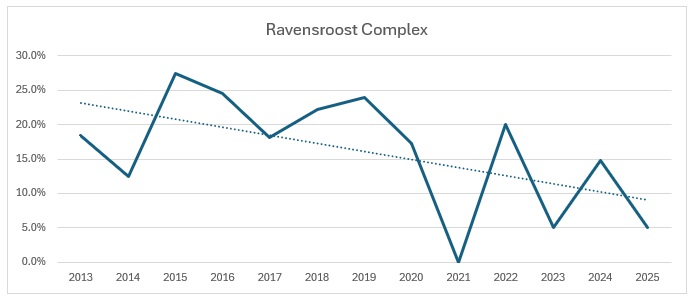
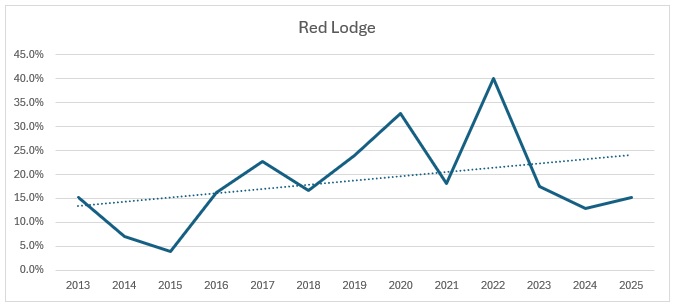
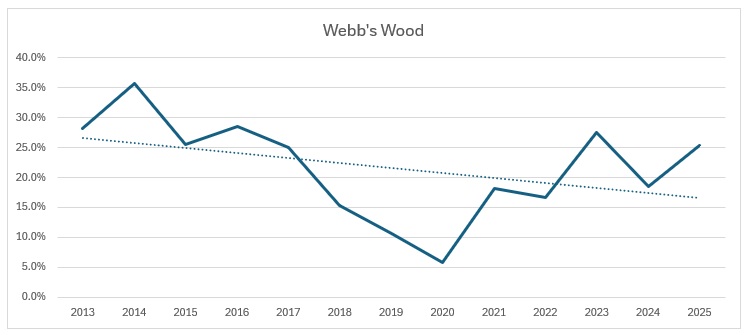
Far too busy to give an indication of where the decline might be, so I have split each out and produced trendlines for each site:
As you can see, the biggest decline is in the Ravensroost complex. followed by Webb’s Wood and the Firs. It just so happens that these three woodlands have all been undergoing significant forestry management over the years. Although Red Lodge did undergo some thinning operations a few years ago, it was in the eastern end of the wood and we work in the western end of the wood. Certainly it didn’t negatively affect the trend, which is a rise of 14% over the period, despite some very up and down results. Somerford Common is clearly the strongest site, also with a 14% rise in numbers over that period.
I am now going to have a look at juvenile recruitment. There is a caveat, of all four of the Paridae species, the Coal Tit is the most difficult to age. You look at the greater coverts and look at the outer fringing of the feathers: if they are blue grey it is adult, if it is yellowish it is juvenile. There is also a colour difference on the main feather body, but it isn’t always clear, the adult feathers being greyer, with a clear white square at the bottom right corner of the feather, when the bird is head away from you, whereas the juvenile feathers are generally duller and browner.
The numbers:
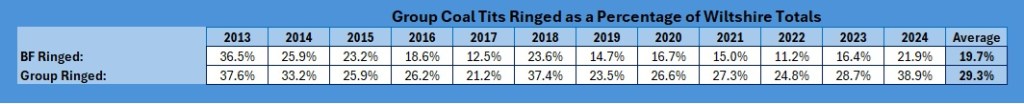
As you can see, the trend across Wiltshire is up and down but the trend is still positive, with an increase of 1% over the 12 years of the study. Our group results are looking stable but those of the Braydon Forest have decreased to negligible.
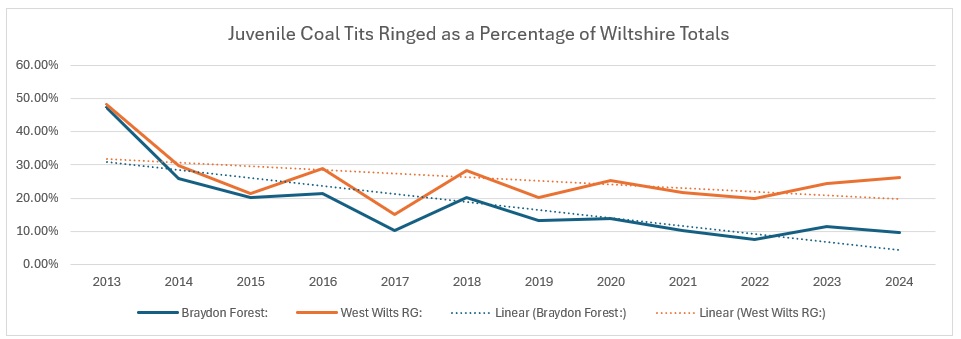
When looked at as a function of the Wiltshire numbers
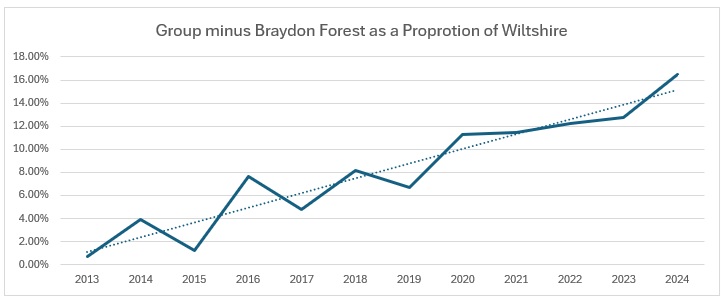
When you look at the trend for the Braydon Forest against the Wiltshire catch, it make for even more depressing reading: from 30% of the Wiltshire total to just 4%. The group contribution, clearly affected by the decline in the Braydon Forest population, is not so bad, with a 10% decline from 30% to 20%.
I then took out the Braydon Forest from our group results and the numbers look like this:
So there has been positive growth in the group numbers: still very small, which is why the impact is so pronounced. When the group’s activity is looked at as a proportion of the Wiltshire catch, it looks like a smooth increase, unlike the bald figures:
This increase in numbers is down to Jonny and Andy from our Group: Jonny taking on the Wiltshire Wildlife Trust reserves at Biss Wood and Green Lane Wood just outside Trowbridge in 2020, although their numbers had fallen away in 2024 and have done so again in 2025. In addition, Andy has caught good numbers at one of his Warminster sites in 2018, 2019 and, not as big as the other two, but a reasonable number in 2024. None of our numbers for the species are exceptional but there are a wide number of contributing sites, but not in any great numbers.
The worrying piece is why the decline in juvenile recruitment in the Braydon Forest has been so pronounced. If we can establish what that is we might be able to do something about it.
Following on from the analysis of Blue and Marsh Tit numbers, I have had a look at the same data for the Great Tit: another species that, along with Blue Tits, are supposed to be being advantaged by garden and supplementary feeding in the winter to the detriment of other species. How do the statistics stand up to scrutiny on that?
As with the Blue Tits, I only have data up to the end of 2024 for the entirety of England and Wiltshire, but have comparative data for my Braydon Forest sites against the entirety of the West Wilts Ringing Group. These are the basic figures:
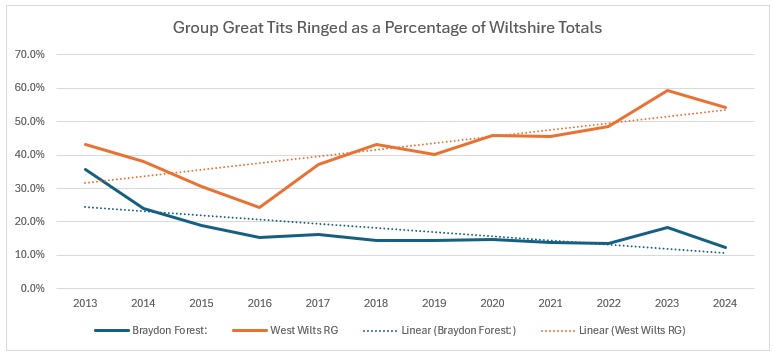
It is interesting to see how small a proportion of the Great Tit catch is found in the Braydon Forest:
The long term trend within the Braydon Forest is a long slow decline: ironically mirroring the trend in Blue Tits. The highs and lows within the Forest are quite smooth, but the growth in the rest of the Group’s activity is significantly different to what is happening there.
How do they stack up against the rest of Wiltshire:
Obviously, only going as far as 2024 does not reflect the rather large hike in the Group numbers in 2025, however, the downward trend in the Braydon Forest is even more pronounced.
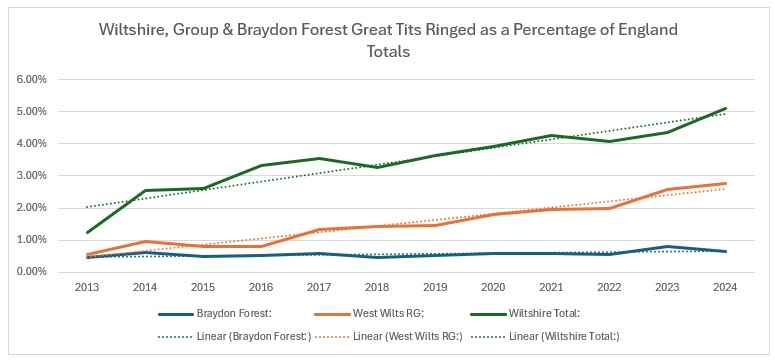
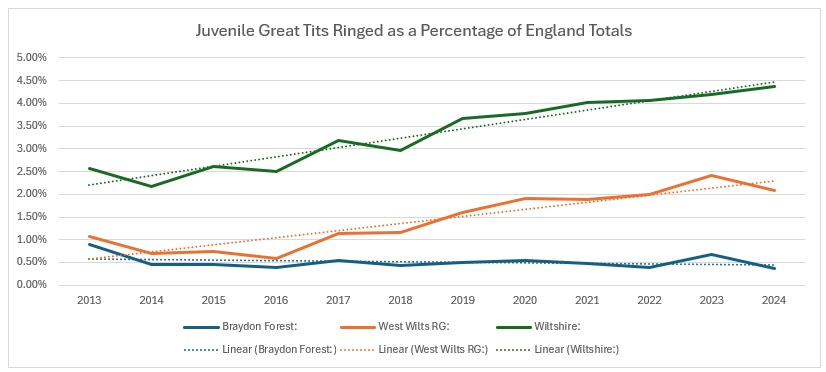
When we compare the Wiltshire trends across the whole of England, the Wiltshire and Group trends are showing a definite increase and the Braydon Forest is neutral.
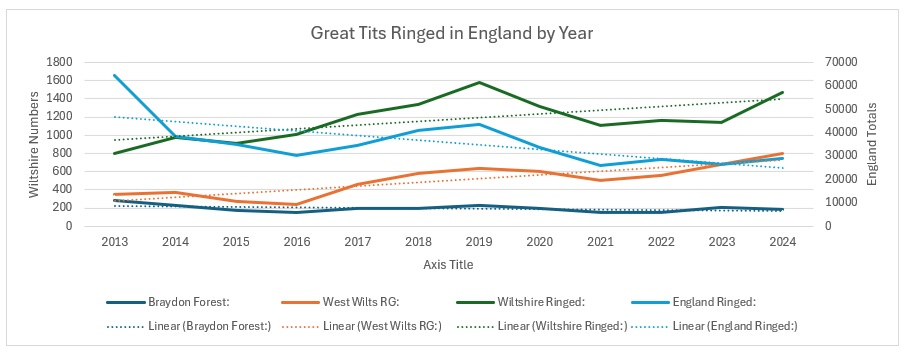
The explanation for that is shown if you graph the actual figures:
As you can see from this, there is a pronounced downward trend of the number ringed across England, even more so than in the Braydon Forest. So it seems that Wiltshire is actually bucking the national trend, with little help from my sites.
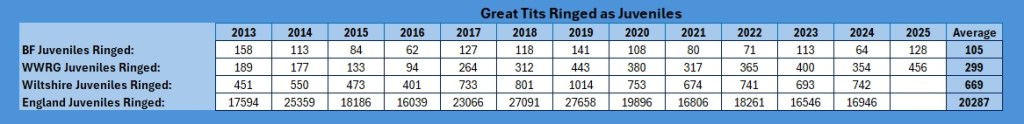
As with the other species, I cannot analyse individuals caught for the whole of Wiltshire and England, as that data is not readily available from the BTO (and I would not want to have to work my way through so much data, it is bad enough doing it for our group). I have looked to see if these overall trends are anything to do with juvenile recruitment. As with the Blue and Marsh Tit pieces, the juvenile analysis does not include pulli ringed as within our group only one of the team monitors titmouse nest boxes, and there are none for me to monitor in the Braydon Forest either. So juveniles ringed are birds ringed post-fledging.
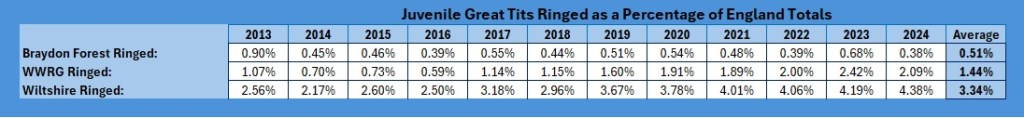
As you can see, the Braydon Forest averages just under one-sixth of the Wiltshire total, the Group as a whole just under half of the Wiltshire total, but when looked at the England total juvenile recruitment, as much as one can extrapolate from ringed birds, the Braydon Forest is producing, on average, 0.51% of the England total, the Group contribution is 1.44% and the whole Wiltshire contribution is just 3.34%.
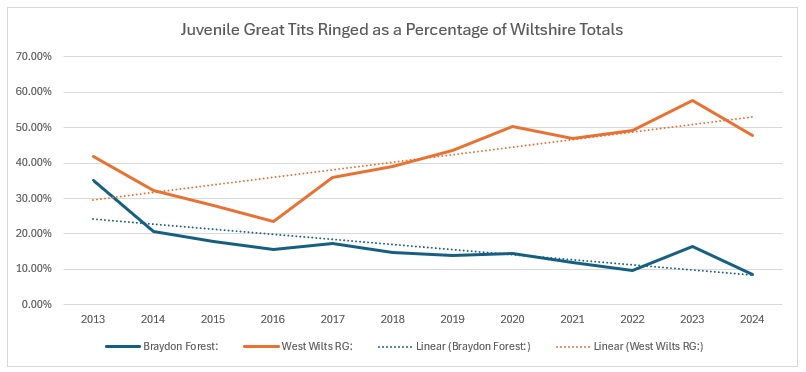
Again, a slight reducing trend within the Braydon Forest, but significant increases with the rest of the group and Wiltshire as a whole.
That reduction is more obvious when you look at the Braydon Forest against the Group and the rest of Wiltshire in graphical form.
There was a slight spike for the Braydon Forest in 2023, but it clearly had little impact on the overall trend.
So, the point of all of this: within the Braydon Forest Marsh Tits are definitely increasing, whereas Blue and Great Tit numbers within it are in decline. There are a number of factors but I think key is the absence of titmouse nest boxes.
We seem to have moved from the Ice Age into the monsoon season in very short order. After our excellent sessions at the Somerford Common sites of the 3rd and 10th of January, the next two sessions had to be cancelled because of the wet weather. Last Wednesday was particularly frustrating: we arrived on site but, although forecast to be at zero, warming through the morning, when we arrived on site it was -7oC and it failed to warm at all. For Saturday it forecast to be wet on three of the usual suspects (Met Office, Meteo and Weather.com) but dry on just one (xcweather). I told the team to have their phones on at 6:30 and I would check the actual weather, as the site is within 2km of my home, and confirm or cancel. I went outside at 6:20 and it was wet, cold and miserable, so I cancelled. Back to bed, woke up at 9:30: sunshine, no wind, lots of swearing!
With this week looking wet all week and beyond, Tuesday was the only available day with a chance of getting out. There were warnings about the wind, but we decided to risk it and see how it turned out. I decided to head to Red Lodge, as we have had some good catches there recently. On Friday I went out and topped up the feeders at every one of my ringing sites. When I arrived on site at 7:15 this morning, the feeders had been emptied, so I was hopeful of a good haul. I topped them up whilst waiting for the team to arrive. The team today comprised Laura, Miranda, Ellie and Pete.
We set the usual nets and tried an extra two, just in case. Suffice to say, we could have not bothered with them: they didn’t catch a single bird!
Given how completely the feeders in ride 3 were emptied in such a short time, I was surprised that it did not start catching until the nets had been open for over an hour. In fact, given that the last two sessions there had produced about 30 birds an hour, today it was a mere 10 birds per hour! It did mean that we didn’t have the hectic sessions that we have had recently. Also, it meant that we weren’t pecked to bits by Blue Tits, so that was one benefit. The downside was that there wasn’t much variety in a relatively small catch.
Not that it was all bad: we had our first Treecreeper of the year:
Treecreeper, Certhia familiaris
We caught and ringed another Marsh Tit: that is three caught in the three Braydon Forest sessions this year! We have also recaptured another nine individual Marsh Tits as well. A good start to the year in the Braydon Forest for this red-listed species.
The list for the morning was: Treecreeper 1; Blue Tit 16(6); Great Tit 3(5) Coal Tit 1(2); Marsh Tit 1(1); Redwing 1. Totals: 23 birds ringed from 9 species and 14 birds retrapped from 4 species, making 37 birds processed from 9 species.
So, not the most exciting catch. The Redwing means that the only woodland site left that needs one for this winter is Ravensroost and, weather permitting, we will be there on Saturday (the forecast does not fill me with confidence).
Unfortunately, at about 10:45 the wind got much stronger and consistently so, so we extracted the last birds and closed the nets. Once we had processed them we took down, packed away and left sit by just after 11:30.